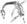Extinction du Capitanien
Intensité des extinctions marines au cours du Phanérozoïque

Le graphique bleu indique le pourcentage apparent (pas en nombre absolu) de genres d'animaux marins ayant disparu au cours d'un intervalle de temps. Il ne représente pas toutes les espèces marines, mais seulement les espèces marines fossiles. Les 5 plus grandes extinctions sont liées, voir les extinctions massives pour plus de détails.
Source et information sur le graphique
L'extinction du Capitanien était un événement d'extinction qui a précédé l'extinction Permien-Trias et s'est produit il y a environ 260 millions d'années pendant une période de diminution de la richesse en espèces et d'augmentation des taux d'extinction à la fin du Permien moyen. Il est également connu sous le nom d'extinction de la fin du Guadalupien en raison de sa reconnaissance initiale entre les séries Guadalupien et du Lopingien ; cependant, une étude stratigraphique plus raffinée suggère que des pics d'extinction dans de nombreux groupes taxonomiques se sont produits au Guadalupien, dans la seconde moitié de l'âge Capitanien[1].
Ampleur

Cette section est vide, insuffisamment détaillée ou incomplète. Votre aide est la bienvenue ! Comment faire ?
Après l'extinction d'Olson, la diversité globale a augmenté au cours du Capitanien. Ce phénomène est probablement dû au remplacement des guildes éteintes par des taxons sinistrés. Bien que les faunes aient commencé à se reconstituer immédiatement après l'extinction capitanienne[2],[3], en reconstruisant des structures trophiques complexes et en remplissant à nouveau les guildes[2], la diversité et la disparité ont continué à diminuer jusqu'à la limite entre le Permien et le Trias[4].
Écosystèmes marins
L'impact de l'extinction du capitanien sur les écosystèmes marins est encore fortement débattu par les paléontologues. Les premières estimations faisaient état d'une perte de genres d'invertébrés marins comprise entre 35 et 47 %[5],[6] tandis qu'une estimation publiée en 2016 suggérait une perte de 33 à 35 % des genres marins après correction pour tenir compte de l'extinction de fond, de l'effet Signor-Lipps et du regroupement des extinctions de certains taxons[7]. La perte d'invertébrés marins au cours de l'extinction de masse du Capitanien était comparable en ampleur à l'événement d'extinction du Crétacé-Paléogène[8].
Certaines études l'ont considérée comme la troisième ou la quatrième extinction massive la plus importante en termes de proportion de genres d'invertébrés marins perdus ; une autre étude a montré que l'extinction capitanienne n'était que la neuvième plus importante en termes de gravité taxonomique (nombre de genres perdus), mais qu'elle était la cinquième plus importante en termes d'impact écologique, c'est-à-dire le degré de restructuration taxonomique dans les écosystèmes ou la perte de niches écologiques, voire d'écosystèmes entiers eux-mêmes[9].
Écosystèmes terrestres
Il existe peu d'estimations publiées concernant l'impact sur les écosystèmes terrestres de l'extinction massive capitanienne. Chez les vertébrés, Day et ses collègues ont suggéré une perte de 74 à 80 % de la richesse générique chez les tétrapodes du bassin de Karoo en Afrique du Sud[10], y compris l'extinction des dinocéphales[11]. Chez les plantes terrestres, Stevens et ses collègues ont constaté une extinction de 56 % des espèces végétales enregistrées dans la formation de Shihhotse supérieure moyenne en Chine du Nord[12], qui était approximativement d'âge mi-capitanien. 24 % des espèces végétales du sud de la Chine se sont éteintes[13].
Chronologie
Cette section est vide, insuffisamment détaillée ou incomplète. Votre aide est la bienvenue ! Comment faire ?
Selon une étude, l'extinction massive du Capitanien n'était pas un événement discret mais un déclin continu de la diversité qui a commencé à la fin du Wordien[14] Une autre étude examinant les faciès fossilifères du Svalbard n'a trouvé aucune preuve d'une extinction massive soudaine, attribuant plutôt les changements biotiques locaux pendant le Capitanien à la migration vers le sud de nombreux taxons à travers la mer de Zechstein[15]. Les dépôts de plates-formes carbonatées de Hongrie et d'Hydra ne montrent aucun signe d'extinction à la fin du Capitanien ; l'extinction y est enregistrée au milieu du Capitanien[16].
Les roches volcaniques des trappes d'Emeishan, qui sont interstratifiées avec les plates-formes carbonatées tropicales de la formation Maokou, sont uniques pour la préservation d'une extinction de masse et la cause de cette extinction de masse[13]. De grandes éruptions phréatomagmatiques se sont produites lorsque les trappes d'Emeishan ont commencé à entrer en éruption, entraînant l'extinction des foraminifères fusulinacés et des algues calcaires[17].
En l'absence d'âges radiométriques contraignant directement les horizons d'extinction eux-mêmes dans les sections marines, la plupart des études récentes s'abstiennent de chiffrer son âge, mais sur la base d'extrapolations de l'échelle de temps du Permien, un âge d'environ 260-262 Ma a été estimé [10],[18], ce qui correspond largement aux âges radiométriques du domaine terrestre, en supposant que les deux événements soient contemporains. Les pertes végétales se sont produites soit en même temps que l'extinction marine, soit après celle-ci[13].
Domaine marin
L'extinction des foraminifères fusulinacés dans le sud-ouest de la Chine était initialement datée de la fin du Guadalupien, mais des études publiées en 2009 et 2010 ont daté l'extinction de ces fusulinacés au milieu du Capitanien[13]. Les pertes de brachiopodes et de coraux se sont produites au milieu du stade Capitanien[19]. L'extinction subie par les ammonoïdes pourrait s'être produite au début du Wuchiapingien[19].
Domaine terrestre
L'existence d'un changement dans les faunes de tétrapodes au milieu du Permien est connue depuis longtemps en Afrique du Sud et en Russie. En Russie, cela correspondait à la limite entre ce qui a été appelé la superzone Titanophoneus et la superzone Scutosaurus[20] et plus tard, respectivement le superassemblage Dinocephalien et le superassemblage Theriodontien. En Afrique du Sud, cela correspondait à la limite entre la zone d'assemblage Pareiasaurus, Dinocephalian ou Tapinocephalus, diversement nommée, et les assemblages sus-jacents[21],[22],[23],[24] En Russie comme en Afrique du Sud, cette transition a été associée à l'extinction du groupe précédemment dominant d'amniotes thérapsides, les dinocephaliens, ce qui a conduit à sa désignation ultérieure sous le nom d'extinction dinocephalienne[25]. Les taux d'apparition après l'extinction sont restés faibles dans la zone d'assemblage de Pristerognathus pendant au moins 1 million d'années, ce qui suggère que les écosystèmes du bassin du Karoo se sont rétablis tardivement[26].
Après la reconnaissance d'une extinction massive marine distincte à la fin du Guadalupien, l'extinction des dinocéphales a été considérée comme son corrélat terrestre[8]. Bien qu'il ait été suggéré par la suite que la faune russe d'Ischeevo, considérée comme la plus jeune faune dinocéphale de cette région, était contrainte de se trouver sous le renversement magnétique de l'Illawarra et devait donc se produire au stade wordien, bien avant la fin du Guadalupien[25].
La reconnaissance d'une faune dinocéphale plus jeune en Russie - l'assemblage de tétrapodes du Sundyr[27] et l'obtention d'âges radiométriques bien contraints sur le plan biostratigraphique par la datation uranium-plomb d'un tuf vocanique de la zone de l'Assemblage de Tapinocephalus du bassin de Karoo[10],[28] ont démontré que l'extinction des dinocéphales s'est effectivement produite à la fin du Capitanien, il y a environ 260 millions d'années.
Effets sur la biodiversité
Cette section est vide, insuffisamment détaillée ou incomplète. Votre aide est la bienvenue ! Comment faire ?
Sur la vie marine
Dans les océans, l'extinction capitanienne a entraîné des taux d'extinction élevés parmi les ammonoïdes, les coraux et les algues calcaires qui construisent les récifs, les foraminifères, les bryozoaires et les brachiopode, et a été plus sévère dans les bassins marins restreints que dans les océans ouverts[20]. Elle a été plus grave dans les bassins marins restreints que dans les océans ouverts et semble avoir été particulièrement sélectif pour les taxons des eaux peu profondes qui dépendaient de la photosynthèse ou d'une relation photosymbiotique[21] ; de nombreuses espèces dont la physiologie respiratoire était mal protégée ont également disparu[22],[23]. L'extinction a entraîné un effondrement de la fabrique de carbonate récifal dans les mers peu profondes entourant la Chine méridionale[24],[25].
Les ammonoïdes, qui avaient connu un déclin à long terme pendant une période de 30 millions d'années depuis le Roadien, ont subi une impulsion d'extinction sélective à la fin du Capitanien[26]. 75,6 % des familles de coraux, 77,8 % des genres de coraux et 82,2 % des espèces de coraux qui se trouvaient dans la Chine permienne ont disparu pendant l'extinction massive du Capitanien[8]. Les Verbeekinidae, une famille de grands foraminifères fusulines, se sont éteints[25].
87 % des espèces de brachiopodes trouvées dans la formation de Kapp Starostin, au Spitzberg, ont disparu sur une période de plusieurs dizaines de milliers d'années ; bien que de nouvelles espèces de brachiopodes et de bivalves soient apparues après l'extinction, la position dominante des brachiopodes a été reprise par les bivalves. Le registre fossile du Groenland oriental est semblable à celui du Spitzberg ; les pertes de faune dans le bassin de Sverdrup au Canada sont comparables aux extinctions au Spitzberg et au Groenland oriental, mais la récupération post-extinction qui s'est produite au Spitzberg et au Groenland oriental ne s'est pas produite dans le bassin de Sverdrup[10]. Alors que les brachiopodes rhynchonelliformes représentaient 99,1 % des individus trouvés dans les carbonates tropicaux de l'ouest des États-Unis, de la Chine du Sud et de la Grèce avant l'extinction, les mollusques représentaient 61,2 % des individus trouvés dans des environnements similaires après l'extinction[29]. 87 % des espèces de brachiopodes et 82 % des espèces de foraminifères fusulinacés de la Chine du Sud ont disparu[18].
Bien que grave pour les brachiopodes, l'impact de l'extinction capitanienne sur leur diversité est loin d'être aussi fort que celui de l'extinction de la fin du Permien[30].
Les biomarqueurs indiquent que les algues rouges et les bactéries photoautotrophes ont dominé les communautés microbiennes marines. Des changements importants dans les écosystèmes microbiens ont eu lieu pendant l'extinction massive du Capitanien, bien qu'ils aient été moins importants que ceux associés à l'extinction de la fin du Permien[31].
La plupart des victimes marines de l'extinction étaient soit des espèces endémiques des mers épicontinentales autour de la Pangée qui sont mortes lorsque les mers se sont refermées, soit des espèces dominantes de l'océan Paléotethys[32]. Des preuves provenant de dépôts marins au Japon et à Primorye suggèrent que la vie marine des latitudes moyennes a été affectée plus tôt par l'événement d'extinction que les organismes marins des tropiques[33].
Les paléontologues ne s'entendent pas sur la question de savoir si et dans quelle mesure la latitude a influé sur la probabilité d'extinction des taxons. Alors que certaines études concluent que l'événement d'extinction était régional et limité aux zones tropicales[34], d'autres suggèrent qu'il y avait peu de variation latitudinale dans les schémas d'extinction[35]. Une étude examinant les extinctions de foraminifères en particulier a révélé que la Paléotéthys centrale et occidentale a connu des pertes taxonomiques d'une ampleur plus faible que la Paléotéthys septentrionale et orientale, qui a connu l'ampleur d'extinction la plus élevée. La même étude a montré que l'ampleur globale de l'extinction de Panthalassa était similaire à celle des Paléotéthys centrale et occidentale, mais que l'ampleur de l'extinction des taxons endémiques était élevée[36].
Cette extinction massive a marqué le début de la transition entre les faunes évolutives paléozoïques et modernes[37]. La transition brachiopode-mollusque qui a caractérisé le passage plus large des faunes évolutives paléozoïques aux faunes évolutives modernes a été suggérée comme ayant ses racines dans l'événement d'extinction massive du Capitanien, bien que d'autres recherches aient conclu qu'il pourrait s'agir d'une illusion créée par un biais taphonomique dans les assemblages de fossiles silicifiés, la transition ne commençant qu'après l'extinction plus cataclysmique de la fin du Permien[38].
Après l'extinction massive du Capitanien, des taxons catastrophes tels que Earlandia et Diplosphaerina sont devenus abondants dans ce qui est aujourd'hui la Chine du Sud[39]. Le rétablissement initial des récifs a consisté en des récifs non métazoaires : biohermes algaux et accumulations de récifs algaux-éponges. Cet intervalle de rétablissement initial a été suivi d'un intervalle de récifs dominés par les tubiphytes, lui-même suivi d'un retour de récifs dominés par les métazoaires et les éponges[40]. Dans l'ensemble, le rétablissement des récifs a duré environ 2,5 millions d'années[41].
Sur la vie terrestre
Parmi les vertébrés terrestres, les principales victimes sont les théropsides dinocéphales, qui étaient l'un des éléments les plus courants de la faune tétrapode du Guadalupien ; un seul genre de dino-céphale a survécu à l'extinction capitanienne[8]. La diversité des anomodontes qui vivaient à la fin du Guadalupien a été réduite de moitié par l'extinction massive du Capitanien[42]. Les survivants terrestres de l'extinction du Capitanien pesaient généralement entre 20 et 50 kg et vivaient généralement dans des terriers[8].
Causes
Cette section est vide, insuffisamment détaillée ou incomplète. Votre aide est la bienvenue ! Comment faire ?
Trapps d'Emeishan
On pense que l'extinction, qui a coïncidé avec le début d'une importante excursion négative du δ13C signifiant une grave perturbation du cycle du carbone[43],[13], a été déclenchée par des éruptions de la grande province ignée des pièges d'Emeishan[44],[45],[12], dont les amas de basalte couvrent actuellement une superficie de 250 000 à 500 000 km2, bien que le volume initial des basaltes ait pu être compris entre 500 000 km3 et plus de 1 000 000 km3[46].
L'âge de l'extinction et le dépôt des basaltes d'Emeishan correspondent bien[47],[48] Les récifs et autres sédiments marins intercalés entre les piles de basalte indiquent que le volcanisme d'Emeishan s'est d'abord développé sous l'eau ; les éruptions terrestres de lave ne se sont produites que plus tard dans la période d'activité de la grande province ignée[49]. Ces éruptions auraient libéré de fortes doses de mercure toxique[50] ; l'augmentation des concentrations de mercure coïncide avec l'excursion négative des isotopes du carbone, ce qui indique une cause volcanique commune[44]. L'enrichissement en coronène à la limite Guadaloupien-Lopingien confirme également l'existence d'une activité volcanique massive ; le coronène ne peut se former qu'à des températures extrêmement élevées créées soit par des impacts extraterrestres, soit par un volcanisme massif, la première hypothèse étant exclue en raison de l'absence d'anomalies d'iridium coïncidant avec des anomalies de mercure et de coronène[51].
On pense qu'une grande quantité de dioxyde de carbone et de dioxyde de soufre a été déversée dans la stratosphère des hémisphères nord et sud en raison de la position équatoriale des pièges d'Emeishan, entraînant un refroidissement soudain et un réchauffement à long terme de la planète[17].
Les pièges d'Emeishan ont rejeté entre 130 et 188 tératonnes de dioxyde de carbone au total, à un rythme compris entre 0,08 et 0,25 gigatonne de dioxyde de carbone par an, ce qui les rend responsables d'une augmentation du dioxyde de carbone atmosphérique qui a été à la fois l'une des plus importantes et l'une des plus précipitées de toute l'histoire géologique de la Terre[52]. Le taux d'émission de dioxyde de carbone pendant l'extinction massive capitanienne, bien qu'extrêmement abrupte, était néanmoins nettement plus lent que celui de l'extinction de la fin du Permien, au cours de laquelle les niveaux de dioxyde de carbone ont augmenté cinq fois plus vite[53].
D'importantes quantités de méthane libérées par les digues et les filons-couches pénétrant dans les gisements riches en charbon ont été impliquées comme un facteur supplémentaire de réchauffement[54], bien que cette idée ait été remise en question par des études concluant plutôt que l'extinction a été précipitée directement par les pièges d'Emeishan ou par leur interaction avec les carbonates de plate-forme[55],[56],[57]. Les émissions des pièges d'Emeishan peuvent également avoir contribué à l'effondrement du bouclier d'ozone, en exposant la surface de la Terre à un flux considérablement accru de rayonnement solaire à haute fréquence[58].
Anoxie et euxinie
Le réchauffement climatique résultant de l'activité de la grande province ignée a été impliqué dans l'anoxie marin[59]. Deux événements anoxiques, l'OAE-C1 du Capitanien moyen et l'OAE-C2 de la fin du Capitanien, se sont produits grâce à l'activité volcanique de l'Emeishan[60]. Les émissions volcaniques de gaz à effet de serre et le réchauffement climatique ont accru l'altération des continents et l'érosion minérale, ce qui, à son tour, a été considéré comme un facteur favorisant l'euxinie océanique[61].
L'euxinie a pu être encore exacerbée par la lenteur croissante de la circulation océanique résultant du réchauffement d'origine volcanique[62]. La nature hydrothermale initiale des pièges d'Emeishan signifiait que la vie marine locale autour de la Chine du Sud aurait été particulièrement menacée par l'anoxie due au développement de hyaloclastites dans des bassins restreints et délimités par des failles[49].
L'expansion de l'anoxie océanique a été supposée se produire légèrement avant l'extinction capitanienne elle-même par certaines études, bien qu'il soit probable que la remontée d'eaux anoxiques avant l'extinction massive ait été un phénomène local spécifique à la Chine du Sud[63].
Hypercapnie et acidification
L'océan étant un puits de carbone qui absorbe le dioxyde de carbone atmosphérique, il est probable que les émissions volcaniques excessives de dioxyde de carbone aient entraîné une hypercapnie marine, qui aurait agi en conjonction avec d'autres mécanismes de destruction pour accroître encore la gravité de la crise biotique[64]. La dissolution du dioxyde de carbone émis par les volcans dans les océans a déclenché une acidification des océans[18],[65], qui a probablement contribué à la disparition de divers organismes marins calcaires, en particulier les bivalves alatoconchides géants[66].
En raison de la plus grande solubilité du dioxyde de carbone dans les eaux froides, l'acidification des océans a été particulièrement meurtrière dans les eaux des hautes latitudes[59]. En outre, les pluies acides auraient été une autre conséquence biocide des émissions intenses de soufre produites par le volcanisme des pièges d'Emeishan[17], ce qui a entraîné l'acidification des sols[18].
Toutefois, certains chercheurs ont mis en doute l'existence d'une acidification significative à l'échelle mondiale, concluant que la perturbation du cycle du carbone était trop faible pour avoir provoqué une baisse importante du pH à l'échelle mondiale[67].
Volcanisme massif
L'analyse des valeurs δ13C et δ18O de l'apatite dentaire des spécimens de Diictodon feliceps du supergroupe Karoo montre une excursion δ13C positive et conclut que la fin du Capitanien a été marquée par une aridification massive dans la région, bien que la température soit restée largement la même, ce qui suggère que le changement climatique mondial n'a pas été à l'origine de l'événement d'extinction[68].
L'analyse des taux d'extinction des vertébrés dans le bassin du Karoo, en particulier dans la formation supérieure d'Abrahamskraal et la formation inférieure de Teekloof, montre que la diminution à grande échelle de la diversité des vertébrés terrestres a coïncidé avec le volcanisme dans les pièges d'Emeishan, bien que des preuves solides d'une relation de cause à effet entre ces deux événements restent insaisissables[69].
Une étude de 2015 a remis en question le fait que l'extinction massive du Capitanien ait été de nature globale ou simplement une crise biotique régionale limitée à la Chine du Sud et à quelques autres régions, ne trouvant aucune preuve d'extinctions terrestres ou marines dans l'est de l'Australie liées aux pièges d'Emeishan ou à tout autre déclencheur d'extinction invoqué pour expliquer la chute de la biodiversité dans les basses latitudes de l'hémisphère Nord[34].
Baisse du niveau de la mer
L'extinction massive du Capitanien a été attribuée à la baisse du niveau de la mer[70], la disparition généralisée des récifs en particulier étant liée à cette régression marine[64]. La limite Guadaloupien-Lopingien a coïncidé avec l'une des régressions marines de premier ordre les plus importantes du Phanérozoïque[71].
Des évaporites et des faciès terrestres recouvrant des dépôts carbonatés marins à travers la transition Guadaloupien-Lopingien prouvent la chute brutale du niveau de la mer à la fin du Guadaloupien[64]. En outre, une énorme discordance est associée à la limite Guadaloupien-Lopingien dans de nombreuses strates à travers le monde[72]. La fermeture de la voie maritime sino-mongole à la fin du Capitanien a été invoquée comme un moteur potentiel de la perte de biodiversité du Paléotéthien[73].
Autres hypothèses
L'assèchement de la planète, la tectonique des plaques et la compétition biologique peuvent également avoir joué un rôle dans l'extinction[71],[12],[74],[68]. Les facteurs d'extinction potentiels proposés comme causes du déclin des récifs à la fin du Guadalupien comprennent les fluctuations de la salinité et les collisions tectoniques des microcontinents[64].
Notes et références
Notes
(en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Capitanian mass extinction event » (voir la liste des auteurs).
Références
- ↑ (en) D. P. G. Bond et al., « The mid-Capitanian (Middle Permian) mass extinction and carbon isotope record of South China », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 292, nos 1-2, , p. 282-294 (DOI 10.1016/j.palaeo.2010.03.056, S2CID 129030843, lire en ligne [PDF])
- ↑ a et b (en) Sarda Sahney et Michael J Benton, « Recovery from the most profound mass extinction of all time », Proceedings of the Royal Society B: Biological Sciences, vol. 275, no 1636, , p. 759–765 (ISSN 0962-8452 et 1471-2954, DOI 10.1098/rspb.2007.1370, lire en ligne, consulté le )
- ↑ (en) Sindbad Zazzali, Sylvie Crasquin, Jean-François Deconinck et Qinglai Feng, « Biodiversity across the Guadalupian-Lopingian Boundary: first results on the ostracod (Crustacea) fauna, Chaotian section (Sichuan Province, South China) », Geodiversitas, vol. 37, no 3, , p. 283–313 (ISSN 1280-9659 et 1638-9395, DOI 10.5252/g2015n3a1, lire en ligne, consulté le )
- ↑ (en) Loïc Villier et Dieter Korn, « Morphological Disparity of Ammonoids and the Mark of Permian Mass Extinctions », Science, vol. 306, no 5694, , p. 264–266 (ISSN 0036-8075 et 1095-9203, DOI 10.1126/science.1102127, lire en ligne, consulté le )
- ↑ (en) J. John Sepkoski, « Patterns of Phanerozoic Extinction: a Perspective from Global Data Bases », dans Global Events and Event Stratigraphy in the Phanerozoic: Results of the International Interdisciplinary Cooperation in the IGCP-Project 216 “Global Biological Events in Earth History”, Springer, , 35–51 p. (ISBN 978-3-642-79634-0, DOI 10.1007/978-3-642-79634-0_4, lire en ligne)
- ↑ (en) Richard K. Bambach, Andrew H. Knoll et Steve C. Wang, « Origination, extinction, and mass depletions of marine diversity », Paleobiology, vol. 30, no 4, , p. 522–542 (ISSN 0094-8373 et 1938-5331, DOI 10.1666/0094-8373(2004)030<0522:OEAMDO>2.0.CO;2, lire en ligne, consulté le )
- ↑ Steven M. Stanley, « Estimates of the magnitudes of major marine mass extinctions in earth history », Proceedings of the National Academy of Sciences, vol. 113, no 42, (ISSN 0027-8424 et 1091-6490, DOI 10.1073/pnas.1613094113, lire en ligne, consulté le )
- ↑ a b c d et e G. J. Retallack, C. A. Metzger, T. Greaver et A. H. Jahren, « Middle-Late Permian mass extinction on land », Geological Society of America Bulletin, vol. 118, nos 11-12, , p. 1398–1411 (ISSN 0016-7606 et 1943-2674, DOI 10.1130/b26011.1, lire en ligne, consulté le )
- ↑ George R. McGhee, Peter M. Sheehan, David J. Bottjer et Mary L. Droser, « Ecological ranking of Phanerozoic biodiversity crises: ecological and taxonomic severities are decoupled », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 211, no 3, , p. 289–297 (ISSN 0031-0182, DOI 10.1016/j.palaeo.2004.05.010, lire en ligne, consulté le )
- ↑ a b c et d (en) Michael O. Day, Jahandar Ramezani, Samuel A. Bowring et Peter M. Sadler, « When and how did the terrestrial mid-Permian mass extinction occur? Evidence from the tetrapod record of the Karoo Basin, South Africa », Proceedings of the Royal Society B: Biological Sciences, vol. 282, no 1811, , p. 20150834 (ISSN 0962-8452 et 1471-2954, PMID 26156768, PMCID PMC4528552, DOI 10.1098/rspb.2015.0834, lire en ligne, consulté le )
- ↑ (en) « South Africa's Great Karoo reveals mass extinction », sur ScienceDaily (consulté le )
- ↑ a b et c Liadan G. Stevens, Jason Hilton, David P.G. Bond et Ian J. Glasspool, « Radiation and extinction patterns in Permian floras from North China as indicators for environmental and climate change », Journal of the Geological Society, vol. 168, no 2, , p. 607–619 (ISSN 0016-7649 et 2041-479X, DOI 10.1144/0016-76492010-042, lire en ligne, consulté le )
- ↑ a b c d et e Volcanism, impacts, and mass extinctions: causes and effects, The Geological Society of America, coll. « Special paper », (ISBN 978-0-8137-2505-5)
- ↑ (en) Matthew E. Clapham, Shuzhong Shen et David J. Bottjer, « The double mass extinction revisited: reassessing the severity, selectivity, and causes of the end-Guadalupian biotic crisis (Late Permian) », Paleobiology, vol. 35, no 1, , p. 32–50 (ISSN 0094-8373 et 1938-5331, DOI 10.1666/08033.1, lire en ligne, consulté le )
- ↑ (en) Sangmin Lee, G.R. Shi, Hans A. Nakrem et Jusun Woo, « Mass extinction or extirpation: Permian biotic turnovers in the northwestern margin of Pangea », GSA Bulletin, vol. 134, nos 9-10, , p. 2399–2414 (ISSN 0016-7606 et 1943-2674, DOI 10.1130/B36227.1, lire en ligne, consulté le )
- ↑ (en) P. B. Wignall, D. P. G. Bond, J. Haas et W. Wang, « CAPITANIAN (MIDDLE PERMIAN) MASS EXTINCTION AND RECOVERY IN WESTERN TETHYS: A FOSSIL, FACIES, AND 13C STUDY FROM HUNGARY AND HYDRA ISLAND (GREECE) », PALAIOS, vol. 27, no 2, , p. 78–89 (ISSN 0883-1351, DOI 10.2110/palo.2011.p11-058r, lire en ligne, consulté le )
- ↑ a b et c (en) Paul B. Wignall, Yadong Sun, David P. G. Bond et Gareth Izon, « Volcanism, Mass Extinction, and Carbon Isotope Fluctuations in the Middle Permian of China », Science, vol. 324, no 5931, , p. 1179–1182 (ISSN 0036-8075 et 1095-9203, DOI 10.1126/science.1171956, lire en ligne, consulté le )
- ↑ a b c et d David P.G. Bond, Paul B. Wignall, Michael M. Joachimski et Yadong Sun, « An abrupt extinction in the Middle Permian (Capitanian) of the Boreal Realm (Spitsbergen) and its link to anoxia and acidification », Geological Society of America Bulletin, vol. 127, nos 9-10, , p. 1411–1421 (ISSN 0016-7606 et 1943-2674, DOI 10.1130/b31216.1, lire en ligne, consulté le )
- ↑ a et b David P.G. Bond, Jason Hilton, Paul B. Wignall et Jason R. Ali, « The Middle Permian (Capitanian) mass extinction on land and in the oceans », Earth-Science Reviews, vol. 102, nos 1-2, , p. 100–116 (ISSN 0012-8252, DOI 10.1016/j.earscirev.2010.07.004, lire en ligne, consulté le )
- ↑ a et b A. G. Sennikov et V. K. Golubev, « Sequence of Permian Tetrapod Faunas of Eastern Europe and the Permian–Triassic Ecological Crisis », Paleontological Journal, vol. 51, no 6, , p. 600–611 (ISSN 0031-0301 et 1555-6174, DOI 10.1134/s0031030117060077, lire en ligne, consulté le )
- ↑ a et b James William Kitching, The distribution of the Karroo vertebrate fauna: with special reference to certain genera and the bearing of this distribution on the zoning of the Beaufort beds, Bernard Price Institute for Palaeontological Research, University of the Witwatersrand, coll. « Memoir - Bernard Price Institute for Palaeontological Research, University of the Witwatersrand ; no. 1 », (ISBN 978-0-85494-427-9)
- ↑ a et b Gideon H. Groenewald, Johann Welman et James A. MacEachern, « Vertebrate Burrow Complexes from the Early Triassic Cynognathus Zone (Driekoppen Formation, Beaufort Group) of the Karoo Basin, South Africa », PALAIOS, vol. 16, no 2, , p. 148 (ISSN 0883-1351, DOI 10.2307/3515526, lire en ligne, consulté le )
- ↑ a et b (en) R. Broom, « V.—On the Permian and Triassic Faunas of South Africa », Geological Magazine, vol. 3, no 1, , p. 29–30 (ISSN 0016-7568 et 1469-5081, DOI 10.1017/S001675680012271X, lire en ligne, consulté le )
- ↑ a et b (en) D. M. S. Watson, « II.—The Zones of the Beaufort Beds of the Karroo System in South Africa », Geological Magazine, vol. 1, no 5, , p. 203–208 (ISSN 1469-5081 et 0016-7568, DOI 10.1017/S001675680019675X, lire en ligne, consulté le )
- ↑ a b c et d (en) Spencer G. Lucas, « Timing and magnitude of tetrapod extinctions across the Permo-Triassic boundary », Journal of Asian Earth Sciences, vol. 36, no 6, , p. 491–502 (DOI 10.1016/j.jseaes.2008.11.016, lire en ligne, consulté le )
- ↑ a et b Michael O. Day et Bruce S. Rubidge, « The Late Capitanian Mass Extinction of Terrestrial Vertebrates in the Karoo Basin of South Africa », Frontiers in Earth Science, vol. 9, (ISSN 2296-6463, DOI 10.3389/feart.2021.631198, lire en ligne, consulté le )
- ↑ (en) V. K. Golubev, « Dinocephalian stage in the history of the Permian tetrapod fauna of Eastern Europe », Paleontological Journal, vol. 49, no 12, , p. 1346–1352 (ISSN 0031-0301 et 1555-6174, DOI 10.1134/S0031030115120059, lire en ligne, consulté le )
- ↑ B. S. Rubidge, D. H. Erwin, J. Ramezani et S. A. Bowring, « High-precision temporal calibration of Late Permian vertebrate biostratigraphy: U-Pb zircon constraints from the Karoo Supergroup, South Africa », Geology, vol. 41, no 3, , p. 363–366 (ISSN 0091-7613 et 1943-2682, DOI 10.1130/g33622.1, lire en ligne, consulté le )
- ↑ Matthew E. Clapham, Shuzhong Shen et David J. Bottjer, « The double mass extinction revisited: reassessing the severity, selectivity, and causes of the end-Guadalupian biotic crisis (Late Permian) », Paleobiology, vol. 35, no 1, , p. 32–50 (ISSN 0094-8373 et 1938-5331, DOI 10.1666/08033.1, lire en ligne, consulté le )
- ↑ (en) Shu-Zhong Shen et Yi-Chun Zhang, « Earliest Wuchiapingian (Lopingian, late Permian) brachiopods in southern Hunan, South China: implications for the pre-Lopingian crisis and onset of Lopingian recovery/radiation », Journal of Paleontology, vol. 82, no 5, , p. 924–937 (ISSN 0022-3360 et 1937-2337, DOI 10.1666/07-118.1, lire en ligne, consulté le )
- ↑ (en) Shucheng Xie, Thomas J. Algeo, Wenfeng Zhou et Xiaoyan Ruan, « Contrasting microbial community changes during mass extinctions at the Middle/Late Permian and Permian/Triassic boundaries », Earth and Planetary Science Letters, vol. 460, , p. 180–191 (DOI 10.1016/j.epsl.2016.12.015, lire en ligne, consulté le )
- ↑ « Pelagic ecosystem recovery after end-Permian mass extinction », sur AccessScience (consulté le )
- ↑ (en) Tomomi Kani, Yukio Isozaki, Ryutaro Hayashi et Yuri Zakharov, « Middle Permian (Capitanian) seawater 87 Sr/ 86 Sr minimum coincided with disappearance of tropical biota and reef collapse in NE Japan and Primorye (Far East Russia) », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 499, , p. 13–21 (DOI 10.1016/j.palaeo.2018.03.033, lire en ligne, consulté le )
- ↑ a et b (en) I. Metcalfe, J.L. Crowley, R.S. Nicoll et M. Schmitz, « High-precision U-Pb CA-TIMS calibration of Middle Permian to Lower Triassic sequences, mass extinction and extreme climate-change in eastern Australian Gondwana », Gondwana Research, vol. 28, no 1, , p. 61–81 (DOI 10.1016/j.gr.2014.09.002, lire en ligne, consulté le )
- ↑ (en) Bethany J. Allen, Matthew E. Clapham, Erin E. Saupe et Paul B. Wignall, « Estimating spatial variation in origination and extinction in deep time: a case study using the Permian–Triassic marine invertebrate fossil record », Paleobiology, vol. 49, no 3, , p. 509–526 (ISSN 0094-8373 et 1938-5331, DOI 10.1017/pab.2023.1, lire en ligne, consulté le )
- ↑ (en) David P. G. Bond et Paul B. Wignall, « Latitudinal selectivity of foraminifer extinctions during the late Guadalupian crisis », Paleobiology, vol. 35, no 4, , p. 465–483 (ISSN 0094-8373 et 1938-5331, DOI 10.1666/0094-8373-35.4.465, lire en ligne, consulté le )
- ↑ (en) R. De la Horra, A.B. Galán-Abellán, J. López-Gómez et N.D. Sheldon, « Paleoecological and paleoenvironmental changes during the continental Middle–Late Permian transition at the SE Iberian Ranges, Spain », Global and Planetary Change, vol. 94-95, , p. 46–61 (DOI 10.1016/j.gloplacha.2012.06.008, lire en ligne, consulté le )
- ↑ (en) Matthew E. Clapham, « Ecological consequences of the Guadalupian extinction and its role in the brachiopod-mollusk transition », Paleobiology, vol. 41, no 2, , p. 266–279 (ISSN 0094-8373 et 1938-5331, DOI 10.1017/pab.2014.15, lire en ligne, consulté le )
- ↑ D. P. G. Bond, P. B. Wignall, W. Wang et G. Izon, « The mid-Capitanian (Middle Permian) mass extinction and carbon isotope record of South China », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 292, no 1, , p. 282–294 (ISSN 0031-0182, DOI 10.1016/j.palaeo.2010.03.056, lire en ligne, consulté le )
- ↑ (en) X. Wang, W.J. Foster, J. Yan et A. Li, « Delayed recovery of metazoan reefs on the Laibin-Heshan platform margin following the Middle Permian (Capitanian) mass extinction », Global and Planetary Change, vol. 180, , p. 1–15 (DOI 10.1016/j.gloplacha.2019.05.005, lire en ligne, consulté le )
- ↑ (en) Yuangeng Huang, Zhong-Qiang Chen, Laishi Zhao et George D. Stanley, « Restoration of reef ecosystems following the Guadalupian–Lopingian boundary mass extinction: Evidence from the Laibin area, South China », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 519, , p. 8–22 (DOI 10.1016/j.palaeo.2017.08.027, lire en ligne, consulté le )
- ↑ (en) Jörg Fröbisch, « Global Taxonomic Diversity of Anomodonts (Tetrapoda, Therapsida) and the Terrestrial Rock Record Across the Permian-Triassic Boundary », PLoS ONE, vol. 3, no 11, , e3733 (ISSN 1932-6203, DOI 10.1371/journal.pone.0003733, lire en ligne, consulté le )
- ↑ (en) Shu-zhong Shen, Chang-qun Cao, Hua Zhang et Samuel A. Bowring, « High-resolution δ13Ccarb chemostratigraphy from latest Guadalupian through earliest Triassic in South China and Iran », Earth and Planetary Science Letters, vol. 375, , p. 156–165 (DOI 10.1016/j.epsl.2013.05.020, lire en ligne, consulté le )
- ↑ a et b (en) Kunyue Ling, Hanjie Wen, Stephen E. Grasby et Haonan Zhao, « The Emeishan large igneous province eruption triggered coastal perturbations and the Capitanian mass extinction: Insights from mercury in Permian bauxite beds », Chemical Geology, vol. 617, , p. 121243 (DOI 10.1016/j.chemgeo.2022.121243, lire en ligne, consulté le )
- ↑ (en) Sakineh Arefifard et Jonathan L. Payne, « End-Guadalupian extinction of larger fusulinids in central Iran and implications for the global biotic crisis », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 550, , p. 109743 (DOI 10.1016/j.palaeo.2020.109743, lire en ligne, consulté le )
- ↑ Volcanism, impacts, and mass extinctions: causes and effects, The Geological Society of America, coll. « Special paper », (ISBN 978-0-8137-2505-5), p. 38
- ↑ (en) Bin He, Yi-Gang Xu, Xiao-Long Huang et Zhen-Yu Luo, « Age and duration of the Emeishan flood volcanism, SW China: Geochemistry and SHRIMP zircon U–Pb dating of silicic ignimbrites, post-volcanic Xuanwei Formation and clay tuff at the Chaotian section », Earth and Planetary Science Letters, vol. 255, nos 3-4, , p. 306–323 (DOI 10.1016/j.epsl.2006.12.021, lire en ligne, consulté le )
- ↑ (en) Hu Huang, Peter A. Cawood, Ming-Cai Hou et Jiang-Hai Yang, « Silicic ash beds bracket Emeishan Large Igneous province to < 1 m.y. at ~ 260 Ma », Lithos, vol. 264, , p. 17–27 (DOI 10.1016/j.lithos.2016.08.013, lire en ligne, consulté le )
- ↑ a et b (en) Dougal A. Jerram, Mike Widdowson, Paul B. Wignall et Yadong Sun, « Submarine palaeoenvironments during Emeishan flood basalt volcanism, SW China: Implications for plume–lithosphere interaction during the Capitanian, Middle Permian (‘end Guadalupian’) extinction event », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 441, , p. 65–73 (DOI 10.1016/j.palaeo.2015.06.009, lire en ligne, consulté le )
- ↑ (en) Stephen E. Grasby, Benoit Beauchamp, David P.G. Bond et Paul B. Wignall, « Mercury anomalies associated with three extinction events (Capitanian Crisis, Latest Permian Extinction and the Smithian/Spathian Extinction) in NW Pangea », Geological Magazine, vol. 153, no 2, , p. 285–297 (ISSN 0016-7568 et 1469-5081, DOI 10.1017/S0016756815000436, lire en ligne, consulté le )
- ↑ (en) Kunio Kaiho, Stephen E. Grasby et Zhong-Qiang Chen, « High-temperature combustion event spanning the Guadalupian−Lopingian boundary terminated by soil erosion », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 618, , p. 111518 (DOI 10.1016/j.palaeo.2023.111518, lire en ligne, consulté le )
- ↑ (en) Qiang Jiang, Fred Jourdan, Hugo K. H. Olierook et Renaud E. Merle, « Volume and rate of volcanic CO 2 emissions governed the severity of past environmental crises », Proceedings of the National Academy of Sciences, vol. 119, no 31, (ISSN 0027-8424 et 1091-6490, DOI 10.1073/pnas.2202039119, lire en ligne, consulté le )
- ↑ (en) Wen-qian Wang, Feifei Zhang, Shuang Zhang et Ying Cui, « Ecosystem responses of two Permian biocrises modulated by CO2 emission rates », Earth and Planetary Science Letters, vol. 602, , p. 117940 (DOI 10.1016/j.epsl.2022.117940, lire en ligne, consulté le )
- ↑ (en) Gregory J. Retallack et A. Hope Jahren, « Methane Release from Igneous Intrusion of Coal during Late Permian Extinction Events », The Journal of Geology, vol. 116, no 1, , p. 1–20 (ISSN 0022-1376 et 1537-5269, DOI 10.1086/524120, lire en ligne, consulté le )
- ↑ (en) Nathan D. Sheldon, Ramananda Chakrabarti, Gregory J. Retallack et Roger M. H. Smith, « Contrasting geochemical signatures on land from the Middle and Late Permian extinction events », Sedimentology, vol. 61, no 6, , p. 1812–1829 (ISSN 0037-0746 et 1365-3091, DOI 10.1111/sed.12117, lire en ligne, consulté le )
- ↑ (en) Jiang Zhu, Zhaochong Zhang, M. Santosh et Shucheng Tan, « Recycled carbon degassed from the Emeishan plume as the potential driver for the major end-Guadalupian carbon cycle perturbations », Geoscience Frontiers, vol. 12, no 4, , p. 101140 (DOI 10.1016/j.gsf.2021.101140, lire en ligne, consulté le )
- ↑ (en) Clément Ganino et Nicholas T. Arndt, « Climate changes caused by degassing of sediments during the emplacement of large igneous provinces », Geology, vol. 37, no 4, , p. 323–326 (ISSN 1943-2682 et 0091-7613, DOI 10.1130/G25325A.1, lire en ligne, consulté le )
- ↑ P. B. Wignall, The worst of times: how life on earth survived eighty million years of extinctions, Princeton University Press, (ISBN 978-0-691-14209-8), p.38
- ↑ a et b (en) David P.G. Bond, Paul B. Wignall et Stephen E. Grasby, « The Capitanian (Guadalupian, Middle Permian) mass extinction in NW Pangea (Borup Fiord, Arctic Canada): A global crisis driven by volcanism and anoxia », GSA Bulletin, vol. 132, nos 5-6, , p. 931–942 (ISSN 0016-7606 et 1943-2674, DOI 10.1130/B35281.1, lire en ligne, consulté le )
- ↑ (en) Huyue Song, Thomas J. Algeo, Haijun Song et Jinnan Tong, « Global oceanic anoxia linked with the Capitanian (Middle Permian) marine mass extinction », Earth and Planetary Science Letters, vol. 610, , p. 118128 (DOI 10.1016/j.epsl.2023.118128, lire en ligne, consulté le )
- ↑ (en) Bolin Zhang, Paul B. Wignall, Suping Yao et Wenxuan Hu, « Collapsed upwelling and intensified euxinia in response to climate warming during the Capitanian (Middle Permian) mass extinction », Gondwana Research, vol. 89, , p. 31–46 (DOI 10.1016/j.gr.2020.09.003, lire en ligne, consulté le )
- ↑ (en) Bolin Zhang, Suping Yao, Wenxuan Hu et Hai Ding, « Development of a high-productivity and anoxic-euxinic condition during the late Guadalupian in the Lower Yangtze region: Implications for the mid-Capitanian extinction event », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 531, , p. 108630 (DOI 10.1016/j.palaeo.2018.01.021, lire en ligne, consulté le )
- ↑ (en) Masafumi Saitoh, Yukio Isozaki, Yuichiro Ueno et Naohiro Yoshida, « Middle–Upper Permian carbon isotope stratigraphy at Chaotian, South China: Pre-extinction multiple upwelling of oxygen-depleted water onto continental shelf », Journal of Asian Earth Sciences, vol. 67-68, , p. 51–62 (DOI 10.1016/j.jseaes.2013.02.009, lire en ligne, consulté le )
- ↑ a b c et d (en) Oliver Weidlich, « Permian reefs re-examined: extrinsic control mechanisms of gradual and abrupt changes during 40 my of reef evolution », Geobios, vol. 35, , p. 287–294 (DOI 10.1016/S0016-6995(02)00066-9, lire en ligne, consulté le )
- ↑ Eric Hand, « Sixth extinction, rivaling that of the dinosaurs, should join the big five, scientists say », Science, (ISSN 0036-8075 et 1095-9203, DOI 10.1126/science.aab2504, lire en ligne, consulté le )
- ↑ (en) Fayao Chen, Wuqiang Xue, Jiaxin Yan et Qi Meng, « The implications of the giant bivalve family Alatoconchidae for the end‐Guadalupian (Middle Permian) extinction event », Geological Journal, vol. 56, no 12, , p. 6073–6087 (ISSN 0072-1050 et 1099-1034, DOI 10.1002/gj.4151, lire en ligne, consulté le )
- ↑ (en) Adam B. Jost, Roland Mundil, Bin He et Shaun T. Brown, « Constraining the cause of the end-Guadalupian extinction with coupled records of carbon and calcium isotopes », Earth and Planetary Science Letters, vol. 396, , p. 201–212 (DOI 10.1016/j.epsl.2014.04.014, lire en ligne, consulté le )
- ↑ a et b (en) Kévin Rey, Michael O. Day, Romain Amiot et Jean Goedert, « Stable isotope record implicates aridification without warming during the late Capitanian mass extinction », Gondwana Research, vol. 59, , p. 1–8 (DOI 10.1016/j.gr.2018.02.017, lire en ligne, consulté le )
- ↑ Michael O. Day et Bruce S. Rubidge, « The Late Capitanian Mass Extinction of Terrestrial Vertebrates in the Karoo Basin of South Africa », Frontiers in Earth Science, vol. 9, (ISSN 2296-6463, DOI 10.3389/feart.2021.631198, lire en ligne, consulté le )
- ↑ A Hallam, « Mass extinctions and sea-level changes », Earth-Science Reviews, vol. 48, no 4, , p. 217–250 (DOI 10.1016/S0012-8252(99)00055-0, lire en ligne, consulté le )
- ↑ a et b (en) D.P.G. Bond, P.B. Wignall, W. Wang et G. Izon, « The mid-Capitanian (Middle Permian) mass extinction and carbon isotope record of South China », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 292, nos 1-2, , p. 282–294 (DOI 10.1016/j.palaeo.2010.03.056, lire en ligne, consulté le )
- ↑ (en) Masafumi Saitoh, Yukio Isozaki, Jianxin Yao et Zhansheng Ji, « The appearance of an oxygen-depleted condition on the Capitanian disphotic slope/basin in South China: Middle–Upper Permian stratigraphy at Chaotian in northern Sichuan », Global and Planetary Change, vol. 105, , p. 180–192 (DOI 10.1016/j.gloplacha.2012.01.002, lire en ligne, consulté le )
- ↑ (en) Shen Shu-Zhong et G. R. Shi, « Paleobiogeographical extinction patterns of Permian brachiopods in the Asian–western Pacific region », Paleobiology, vol. 28, no 4, , p. 449–463 (ISSN 0094-8373 et 1938-5331, DOI 10.1666/0094-8373(2002)028<0449:PEPOPB>2.0.CO;2, lire en ligne, consulté le )
- ↑ (en) Xiang‐Dong Wang et Tetsuo Sugiyama, « Diversity and extinction patterns of Permian coral faunas of China », Lethaia, vol. 33, no 4, , p. 285–294 (ISSN 0024-1164 et 1502-3931, DOI 10.1080/002411600750053853, lire en ligne, consulté le )
 Portail de la paléontologie
Portail de la paléontologie  Portail origine et évolution du vivant
Portail origine et évolution du vivant